FKBP8 connects the Hsp70-Hsp90 chaperone machinery to the folding of membrane proteins
Date:2026-07-01
Membrane protein biogenesis in the endoplasmic reticulum (ER) requires coordinated folding of transmembrane domains and cytosolic regions within a crowded membrane environment. While cytosolic Hsp70–Hsp90 chaperone systems are known to assist protein folding, how these machineries are spatially organized at the ER membrane to ensure efficient maturation of membrane proteins has remained unclear.
In a study published in Nature Communications, teams led by Prof. Zai-Rong Zhang and Prof. Yixiao Zhang, in collaboration with Dr. Jing Li’s group at Genentech identify FKBP8 as a membrane-anchored co-chaperone that recruits and organizes the Hsp70–Hsp90 chaperone network at the ER surface. This spatial organization enables efficient folding and dimerization of complex membrane proteins, including the ABC transporter ABCG2. The work reveals a previously unrecognized membrane-localized protein quality control pathway essential for membrane protein biogenesis.
Using a genome-wide CRISPR-Cas9 screen, the authors identified FKBP8 as a key factor required for the maturation of ABCG2, a clinically relevant multidrug resistance transporter. Loss of FKBP8 led to severe defects in ABCG2 biogenesis: newly synthesized protein failed to form functional dimers, accumulated as misfolded monomers in the ER, and was rapidly eliminated through ER-associated degradation (ERAD), resulting in markedly reduced cell surface expression.
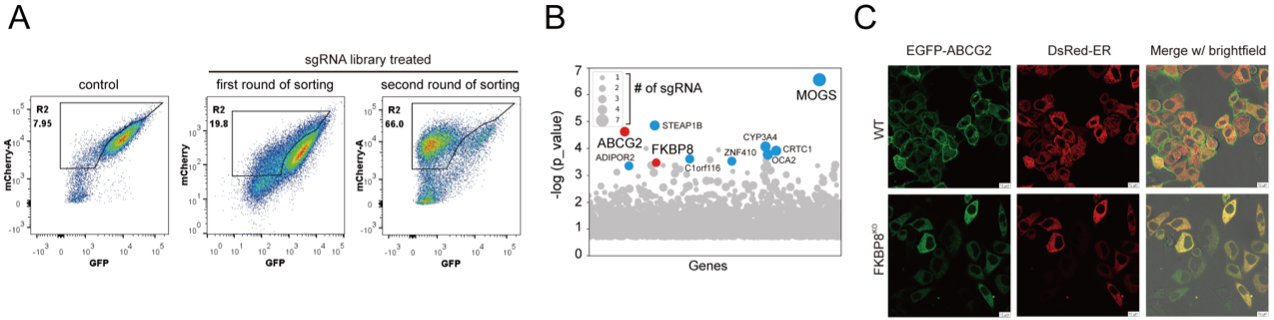
Figure 1. Genome-wide screening identified FKBP8 for the biogenesis of ABCG2
(A) Flow cytometry analysis of GFP fluorescence in control (left) and sgRNA library-treated cells during the first (middle) and second (right) rounds of sorting. The gate “R2” indicates the sorting region, intended to broadly capture cell populations with reduced GFP/mCherry signals. Sequential rounds of sorting resulted in enrichment of these populations.
(B) Analysis of p-values and sgRNA counts for genes identified by deep sequencing. Top hits supported by three or more sgRNAs are highlighted. CRISPR screen data were analyzed using MAGeCK with the robust rank aggregation (RRA) method.
(C) Subcellular localization of ABCG2 in wild-type and FKBP8 KO cells. Wild-type and FKBP8 KO HeLa cells expressing EGFP-tagged ABCG2 and ER-localized DsRed were analyzed by live-cell fluorescence imaging.
Mechanistically, FKBP8 acts independently of its canonical peptidyl-prolyl isomerase (PPIase) activity. Instead, it functions as a membrane-anchored co-chaperone scaffold that recruits Hsp90 and coordinates the Hsp40–Hsp70–HOP chaperone cascade at the ER membrane. This organization positions the cytosolic chaperone machinery directly at sites of membrane protein folding, thereby facilitating maturation of cytosolic domains and subsequent dimerization of ABCG2. Mutational analysis revealed that a conserved N-terminal hydrophobic motif of FKBP8 is critical for substrate engagement, while structural elements within its FKBD domain contribute to stabilizing interactions with client proteins and chaperones.
To elucidate the structural basis of this process, the team determined the cryo-electron microscopy structure of the FKBP8–Hsp90 complex. FKBP8 interacts with the C-terminal region of Hsp90 via its helix 7 extension, forming a stable membrane-associated assembly. This architecture creates a confined chaperone environment at the ER membrane, where FKBP8 and Hsp90 cooperate to coordinate substrate folding. Importantly, Hsp90 ATPase activity is required for this process, indicating that the canonical Hsp70–Hsp90 folding cycle is preserved but spatially redirected to the membrane surface.
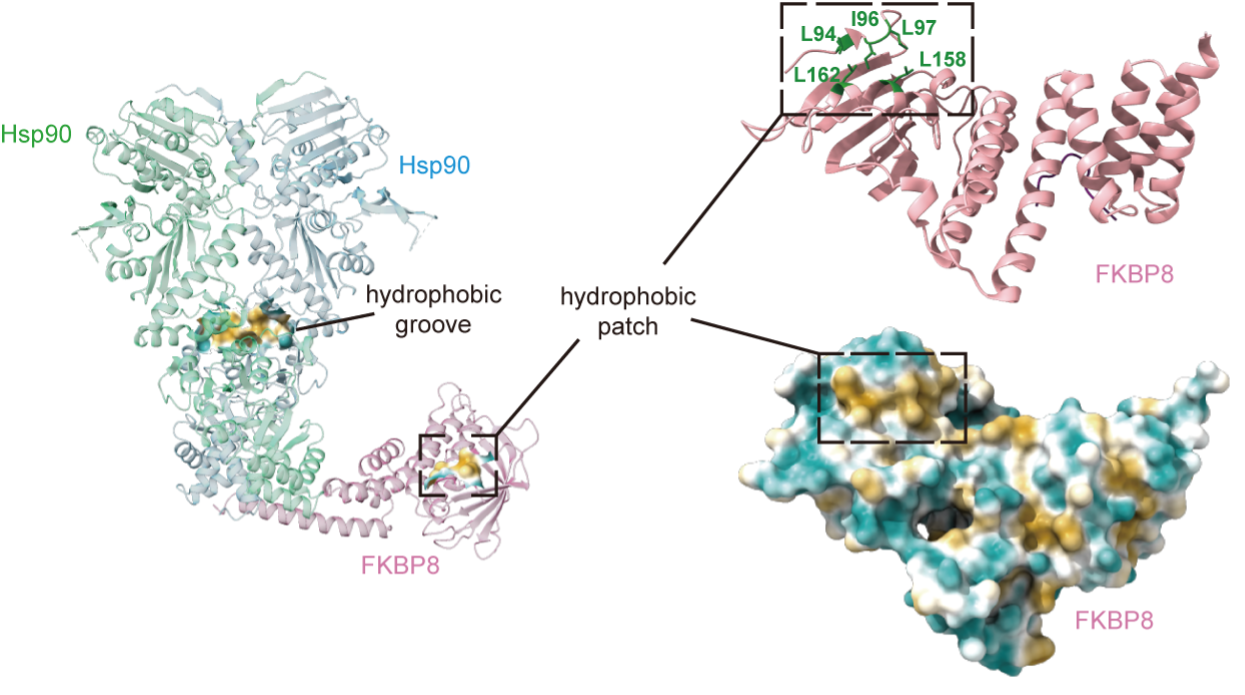
Figure 2. Cryo-EM structure of Hsp90-mFKBP8 complex reveals a substrate-engaging cavity with a hydrophobic cluster required for chaperone activity.
Together, these findings establish FKBP8 as a key organizer of cytosolic chaperone activity at the ER membrane, bridging soluble folding machineries with membrane-localized protein quality control. This spatial coupling provides a mechanistic explanation for how complex membrane proteins achieve efficient folding and assembly in a highly crowded cellular environment.
The study reveals a previously unrecognized principle of membrane protein quality control, in which a membrane-anchored co-chaperone recruits and spatially organizes cytosolic chaperone networks to ensure productive folding. This mechanism expands the classical view of Hsp70–Hsp90 function beyond the cytosol and highlights how spatial regulation enhances proteostasis at the ER membrane.
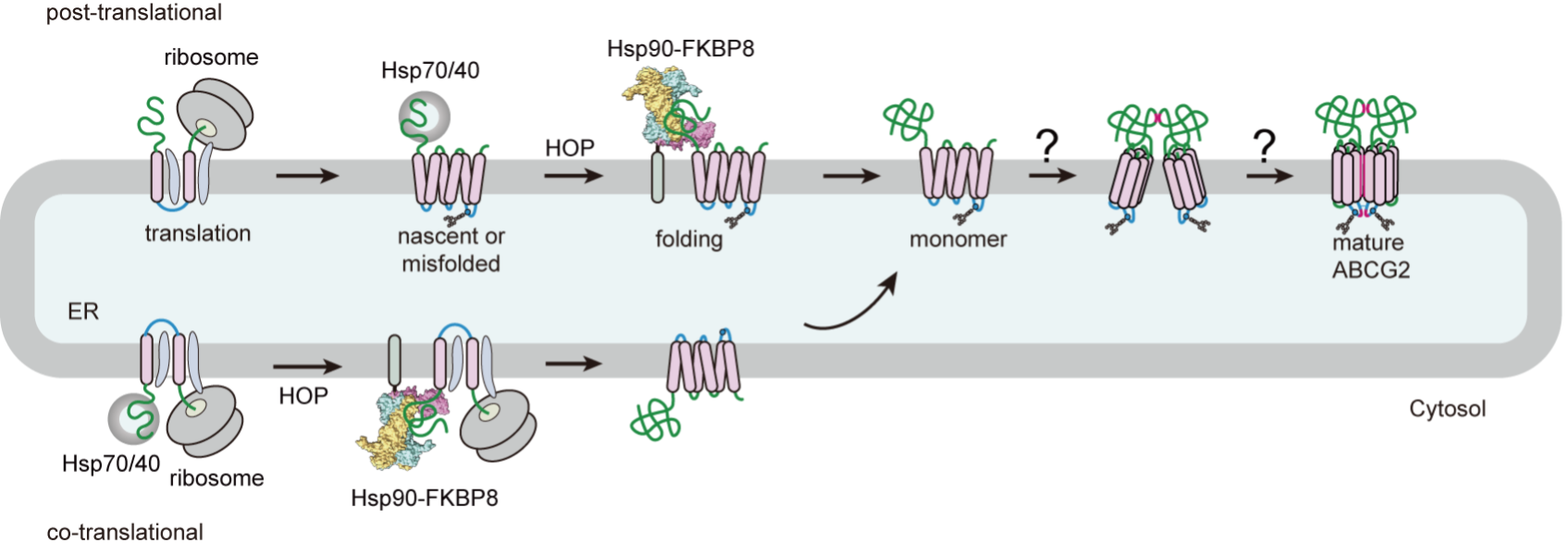
Figure 3. Model of post- and co-translational folding of ABCG2 cytosolic domain by Hsp90-FKBP8 prior to tri-partite assembly between cytosolic, transmembrane, and extracellular domains. Hsp90-FKBP8 mediates folding of nascent and misfolded polypeptide of membrane protein cytosolic domain to facilitate complex assembly.
Given that membrane protein misfolding is associated with cancer drug resistance, neurodegenerative disorders, and genetic diseases, these findings may provide a foundation for developing therapeutic strategies targeting chaperone organization and membrane protein quality control pathways.
This research was supported by the National Natural Science Foundation of China, the Shanghai Municipal Science and Technology Major Project, and the Shanghai Basic Research Pioneer Program.
Article Link: https://www.nature.com/articles/s41467-026-74519-6
附件下载: